| Переменная |
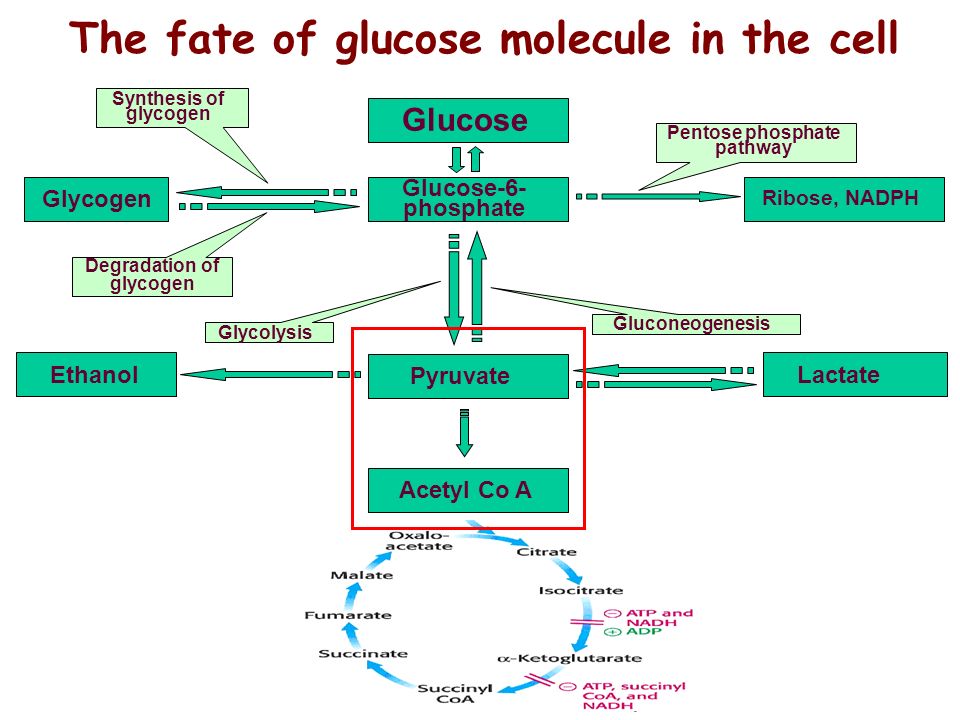
Глюкоза |
Жир/кетоны |
| Реактивные виды кислорода |
Мало при умеренном потреблении;
Много (потенциальный вред митохондриям) при гипергликемии |
Умеренно (вызывает адаптационные изменения) |
| Способность быстро генерировать АТФ |
Да,
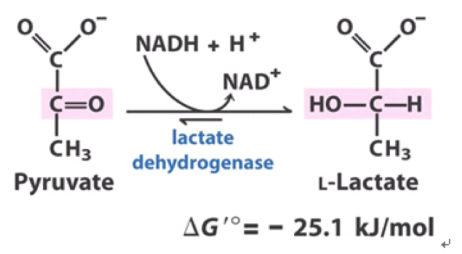
NADH-ориентированный метаболизм через 1й комплекс (2,5 АТФ, выкачка протонов);Пиковая возможность генерировать энергию упираться в доступность NAD+. И скорость получения последнего при помощи восстановления пирувата до лактата. |
Нет,
Есть предел «пиковой бета-оксидации»
Сбалансированный метаболизм NADH/FADH2 1:2, 1:3 (FADH2 дает 1,5 АТФ и не выкачивает протоны) |
| Транспорт в клетки |
Пассивные мембранные транспортеры (GLUT) со специфичной тканям чувствительностью глюкозе;
Ряд GLUT-комплексов требуют присутствия инсулина (например, GLUT4 в мышцах и адипоцитах) |
VLDL;
Кетоны для мозга (VLDL не может пересекать ГЭБ) |
| Способы утилизации |
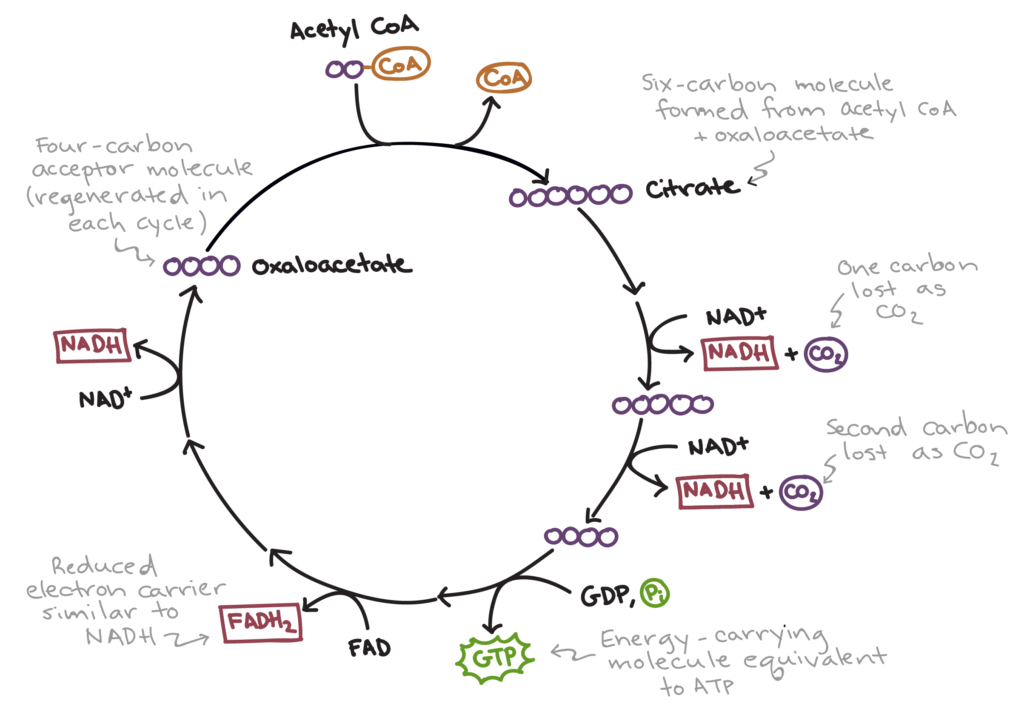
Пируват (белки, цикл Кребса итд);
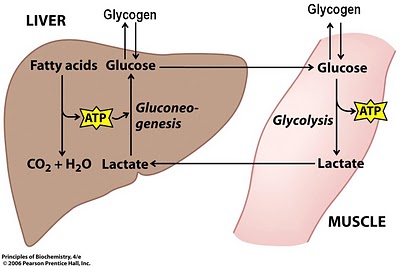
Гликоген;
Пентозо-фосфатный путь (пуриновый метаболизм, NADPH итд) |
Ацетил Ко-А (только на энергию в цикле Кребса)
Синтез жиров и гормонов |
| Последствия переедания |
Лактоацидоз;
Псевдогипоксия;
Гликизирование белков |
Кетоны большом количестве также снижают pH крови (как при диабетическом кетоацидозе), но даже при продолжительном голодании таких показаний сложно добиться. |


 LinkBack URL
LinkBack URL About LinkBacks
About LinkBacks



 Ответить с цитированием
Ответить с цитированием




Социальные закладки